A broader heterobifunctional tool emerges
Targeted protein degradation has rapidly progressed from a conceptual curiosity to one of the most closely watched areas in drug discovery. As discussed in our previous articles in this mini-series, PROTACs have now reached the clinic, with early regulatory milestones signalling their translational potential, while molecular glue degraders continue to demonstrate how subtle manipulation of protein-protein interactions can unlock new therapeutic strategies.
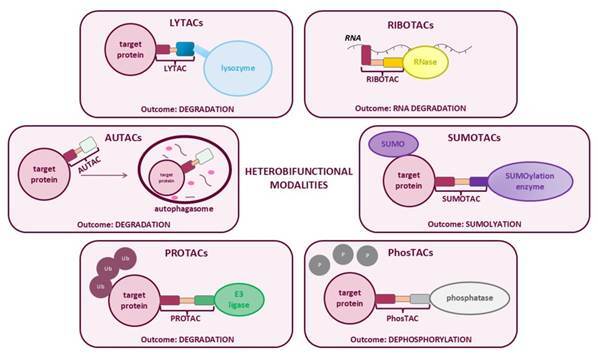
Inspired by these developments, a broader class of heterobifunctional modalities that extend these principles in new directions is rapidly emerging. Technologies such as lysosome-targeting chimeras (LYTACs), autophagy-targeting chimeras (AUTACs), and ribonuclease-targeting chimeras (RIBOTACs) are expanding the degradation paradigm beyond the proteasome. In doing so, they promise access to targets that have historically remained beyond the reach of conventional small molecules, while at the same time raising a fresh set of intellectual property considerations.
A key limitation of both PROTACs and molecular glues is that they rely on intracellular ubiquitin-proteasome pathways. Many therapeutically relevant proteins, including extracellular ligands, membrane-bound receptors, and RNA targets, fall outside this scope. The next generation of degradation technologies seeks to address these gaps by co-opting alternative cellular pathways, a few of which we will outline in this blog.
LYTACs: opening up extracellular and membrane targets
LYTACs represent one of the most conceptually transformative advances in the field. Developed initially in the laboratory of Carolyn Bertozzi, this technology enables targeted degradation of extracellular and membrane proteins by exploiting lysosomal trafficking pathways. Rather than acting within the cytoplasm, LYTACs operate at the cell surface, hijacking endogenous receptor-mediated endocytosis to deliver bound targets to the lysosome for degradation.
In practice, LYTACs combine a target-binding moiety (often an antibody or protein ligand) with a ligand for lysosome-shuttling receptors such as CI-M6PR or ASGPR. This modular design enables selective internalisation and degradation of proteins that have traditionally been addressed using biologics, rather than small molecules.
Commercial interest has quickly followed. Lycia Therapeutics, which emerged from Bertozzi’s work, is a key player advancing LYTAC-based platforms, while other companies are beginning to explore related lysosome-mediated strategies. The modality sits at an interesting interface between small molecules and biologics, and this convergence is also reflected in the IP landscape. Protecting LYTACs may involve not only defining the heterobifunctional constructs themselves, but also navigating overlapping rights in targeting ligands, receptor-binding moieties, and delivery architectures.
AUTACs: harnessing the cell’s recycling machinery
AUTACs take a distinct approach, leveraging the autophagy pathway rather than lysosomal trafficking per se. AUTACs function by tagging target molecules with signals that trigger selective autophagy. Unlike the proteasome, autophagy can degrade larger substrates, including protein aggregates, damaged organelles, and even invading pathogens.
This capability makes AUTACs particularly attractive for disease areas such as neurodegeneration, where the accumulation of toxic aggregates or dysfunctional cellular components is a central feature. Although still at an earlier stage of development, AUTACs are attracting growing attention from both academic groups and emerging biotech companies operating in the broader autophagy and targeted degradation space.
From an IP perspective, AUTACs raise a number of familiar but nuanced issues. A key question is how broadly autophagy-inducing tags or signals can be claimed, particularly where the underlying biology remains relatively complex and not fully characterised. As with earlier degradation technologies, there is likely to be a threshold for robust experimental support demonstrating a clear causal link between construct design and degradation via the intended pathway.
RIBOTACs: extending degradation to RNA
RIBOTACs push the concept of targeted degradation even further, shifting the focus from proteins to RNA. Pioneering work in this area, including by Arrakis Therapeutics and academic collaborators, has demonstrated that small molecules can be designed to recruit endogenous ribonucleases, such as RNase L, to selectively degrade disease-relevant RNA molecules.
This opens up a complementary strategy to established RNA-targeting approaches such as antisense oligonucleotides and RNA interference. Rather than blocking translation or modifying splicing, RIBOTACs aim to directly eliminate the RNA species of interest.
However, moving into RNA targeting introduces additional complexity. RNA structure and folding can play a decisive role in determining accessibility and binding, making target definition less straightforward than for proteins. The IP landscape is also comparatively crowded, with significant prior art relating to RNA therapeutics. As a result, demonstrating inventive step and ensuring sufficient enablement may require particularly careful positioning, supported by detailed structural and functional data.
Beyond degradation: towards programmable modulation
A particularly important trend is the shift beyond degradation towards broader functional modulation. Rather than eliminating targets, newer heterobifunctional approaches are increasingly aimed at reprogramming protein behaviour.
PhosTACs provide a good example of this shift. Mechanistically, they bring a phosphatase enzyme into proximity with a phosphorylated target protein, promoting selective dephosphorylation. This enables precise intervention in signalling pathways without the need to block upstream kinases, potentially offering improved selectivity and fewer off-target effects.
Similarly, emerging SUMOTAC approaches seek to recruit SUMOylation machinery to specific substrates, thereby altering protein stability, localisation or stress responses through installation of SUMO modifications. While still largely at an early stage, this reflects a broader move towards exploiting post‑translational modifications as controlled outputs of induced proximity.
From a commercial standpoint, these approaches represent a significant expansion in mechanistic space. Rather than competing directly with degraders or inhibitors, they open up new therapeutic strategies based on fine-tuning protein function, aligning with a wider industry shift towards more precise and controllable interventions.
|
Modality |
Target class |
Degradation pathway |
Outcome |
|
PROTAC (previous post here) |
Intracellular proteins |
Ubiquitin-proteasome |
Degradation |
|
Molecular glue (previous post here) |
Intracellular proteins (neo-substrates) |
Ubiquitin-proteasome |
Degradation |
|
LYTAC |
Extracellular proteins & membrane receptors |
Lysosomal trafficking |
Degradation |
|
AUTAC |
Aggregates, organelles |
Autophagy |
Degradation |
|
RIBOTAC |
RNA targets (mRNA, non-coding RNA) |
RNase-mediated cleavage |
Degradation |
|
PhosTAC |
Phosphorylated proteins |
Altered signalling |
Functional modulation |
|
SUMOTAC |
Intracellular proteins |
Altered stability, localisation, signalling |
Functional modulation |
IP themes: familiar foundations, new complexity
Despite their differences, these emerging modalities share common strategic considerations. Many of the core IP themes will be familiar from the evolution of PROTACs and molecular glues, including the balance between platform-level claims and target-specific protection, the importance of robust experimental evidence, and the role of comparative data in establishing inventive step.
At the same time, these newer technologies introduce additional layers of complexity. They rely on less well-characterised biological pathways, broaden the scope of addressable targets, and intersect with adjacent modalities such as biologics and RNA therapeutics. Each of these factors can complicate both patent drafting and prosecution, as well as freedom-to-operate analyses.
The field is also being shaped by a mix of academic innovation and commercial activity. Companies such as Lycia Therapeutics and Arrakis Therapeutics, alongside a growing number of early-stage ventures, are helping to define the landscape, while larger pharmaceutical companies are increasingly expanding their interest beyond first-generation degradation technologies.
Looking ahead
These emerging heterobifunctional modalities represent a natural next step in the evolution of targeted degradation. While PROTACs and molecular glues have shown how induced proximity can be exploited to drive protein degradation, LYTACs, AUTACs and RIBOTACs demonstrate that the concept is far more versatile than initially imagined.
For innovators, success will depend not only on advancing the underlying science, but also on securing well-structured and forward-looking IP protection in a landscape that is evolving rapidly and becoming increasingly interconnected. As this second wave of degradation technologies continues to develop, those able to combine scientific innovation with strategic IP positioning are likely to play a defining role in shaping the future of the field.
Do get in touch if you would like to discuss any IP considerations relating to degradation technologies or if you need support to help build a strong IP position in this field.


.png)